|
|
Mecanisme neuroimunologice in SM
Dupa TUTA S. (2002) baza dezvoltarii mijloacelor terapeutice actuale, dezvoltate in deceniul creierului si a altor mijloace terapeutice care sunt in curs de elaborare, o constituie cunoasterea mecanismelor neuroimunologice lezionale din SCLEROZA MULTIPLA. Aceste mecanisme se succed intr-o anumita succesiune. Acestea ar actiona in trei etape:
Etapa I - de prezentare a antigenului;
Etapa II - de traversare de catre limfocitele activate prin bariera hematoencefalica;
Etapa III - nevraxiala de agresiune imunoinflamatorie axomielica.
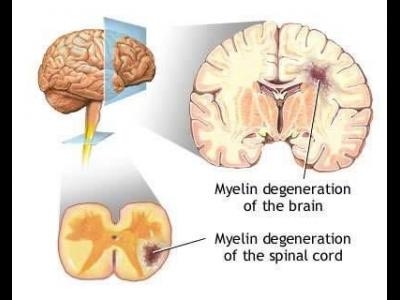
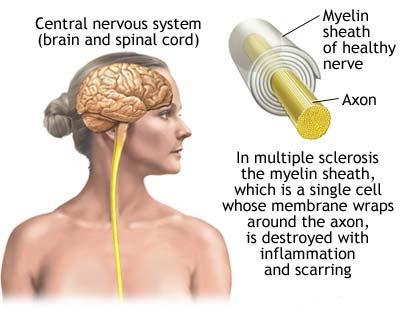
In prima etapa se formeaza molecule proteice care se implica ca antigen inductor in fazele cascadei autoimune din SCLEROZA MULTIPLA. Aceste antigene sunt proteina bazica mielica, proteina proteolipidica, glicoproteina mielica oligodendrocitica, proteina specifica oligodendrocitica si alfa -beta cristalin.
In urma unor agresiuni facute de anumiti virusi sau bacterii asupra sistemului nervos central, iau nastere aceste antigene. Tot antigene sunt si unele fragmente proteice de aminoacizi, apartinatoare acestor virusi si bacterii. Toate aceste antigene sunt preluate de macrofage, microglie, celulele B, in stransa legatura cu moleculele complexului de histocompatibilitate. Acest complex antigenic este recunoscut de un receptor al unor celule T. Aceasta recunostere este primul semnal in reactiile autoimune, care se declanseaza. Al doilea semnal activator activeaza celulele T cu ajutorul unor liganzi. Citochinele au un efect modulator asupra acestui al doilea semnal.
Dupa prezentarea antigenului celulelor T, acestea se diferentiaza in celule efectorii. Aceste celule T cu molecule CD4 pe suprafata, numite si celule T helper se diferentiaza in celule T helper 1 si celule T helper 2. Celulele T helper 1 produc citokine cum ar fi interferonul gama, factorul de necroza tumorala alfa, interleuchina 2 si interleuchina 12, producand activarea macrofagelor. Celulele T helper 2 produc citokinele 1L4, 1L5, 1L10 si factorul de crestere-transformare beta, activand deasemenea si actiunea celulelor B care produc imunoglobuline. Prin urmare celulele T helper 1 au efect proinflamator, iar celulele T helper 2 au un efect antiinflamator si reglator. Raspunsul patologic imun la antigenele specifice create se realizeaza prin doua cai: o cale pe care fragmente proteice virale, asemanatoare fragmentelor proteice mielinice sunt recunoscute ca antigen, cu aparitia unei reactivitati imune incrucisate, si o a doua cale in care activitatea unui factor lezional endogen inflamator, sau exogen (viral, bacterian), produce fragmente peptidice antigenice mielinice, sau ale unor alte componente importante din sistemul nervos central, cum ar fi oligodendrocitele, declansand o multime de reactii autoimune. Toate aceste reactii se produc pe un teren de predispozitie genetica, cu participarea unor factori de mediu, cu scaderea imunitatii date de limfocitele T.
Studiind fenomenele ce apar in etapa a II-a de traversare a barierei hematoencefalice de catre limfocitele active, se poate observa ca in mod normal celulele endoteliale ale capilarelor din sistemul nervos central, sunt impermeabile pentru limfocitele din tesutul sangvin. In timpul raspunsului inflamtor, factorul de necroza tumorala alfa si interferonul gama induce la nivelul celulelor endoteliale fenomene care fac posibila cresterea adezivitatii celulelor T. Astfel apar infiltratele perivasculare in jurul venelor si capilarelor care contin celule T, aceste infiltrate fiind caracteristice SM si EAE.
Aceste infiltrate sunt limitate extern de o matrice extracelulara. Pentru a leza sistemul nervos limfocitele din infiltratele perivasculare trebuie sa treaca bariera matricei extracelulare, care contine fibre de colagen. Neuroproteazele matriceale sunt enzime care degradeaza matricea extracelulara si sunt implicate in proteoliza mielinei in SCLEROZA MULTIPLA. De asemenea, gelatinazele joaca un rol cheie in penetrarea matricei extracelulare de catre limfocitele activate. Aceste gelatinaze sunt detectabile in lichidul cefalo-rahidian, in celulele endoteliale, macrofage si astrocitele bolnavilor cu SCLEROZA MULTIPLA. Gelatinazele produc lezarea colagenului din structura membranei bazale, ajutand la penetrarea barierei hematoencefalice de catre limfocitele activate. Din cercetarile efectuate in timpul administrarii de interferon beta bolnavilor cu SM, s-a observat ca acesta este un puternic inhibitor al activitatii gelatinazelor, limitand patrunderea celulelor T in sistemul nervos central.
Etapa a III-a se petrece in sistemul nervos central si consta in agresiunea imuno-inflamatorie axomielica a anticorpilor secrectati de limfocitele B, impotriva numeroaselor proteine mielice si lipide ale invelisului mielic.
Tintele principale ale anticorpilor sunt proteina bazica mielica, proteina proteolipidica, glicoproteina mielica oligodendrocitara, proteina specifica oligodendrocitara, precum si alfa-beta crystalin, care apare in teaca de mielina dupa declansarea raspunsului inflamator (numita si proteina de stress). Un mediator al agresiunii in afectiunile autoimune este radicalul liber oxid nitric (NO). El este implicat in distructia oligodendrogliei de catre celulele microgliale. Oxidul nitric a fost identificat in leziunile demielinizante din SCLEROZA MULTIPLA. Interferonul gama si factorul de necroza tumorala alfa favorizeaza patrunderea oxidului nitric in microglie, osteocite si macrofage. Anticorpii, oxidul nitric si factorul de necroza tumorala alfa distrug mielina si activeaza macrofagele pentru a fagocita componentele mielinice. Celulele T activate si macrofagele mai produc o substanta numita osteopontin, care mobilizeaza mai multe celule T helper 1 in productia de citokine proinflamatoare de tipul interferon gama si 1L-l2.
Osteopontinul suprima activitatea citokinelor antiinflamatoare produse de celulele T helper 2. Astfel se produce un dezechilibru intre activitatea inflamatorie si distructiva a celulelor T helper 1 si cea antiinflamatorie si reglatoare a celulelor T helper 2, in favoarea primelor. In felul acesta in sistemul nervos central apar zone intinse de demielinizare si leziuni oligodendrogliale.
In timpul inflamatiei din sistemul nervos al bolnavilor cu SM, limfocitele T, microgliile si macrofagele pun in libertate cantitati mari de glutamat, care activeaza receptorii alfa-amino-3hidroxi-5metil-4 isoxazolepropionic acid, mediind (crescand) toxicitatea indusa de neurotransmitatorii excitatori tip glutamat. Leziunile mediate de glutamat, prin intermediul receptorilor alfa-amino-3hidroxi-4metil-4isoxazolepropionic acid se produc printr-un influx ridicat de calciu intracelular, cu leziuni necrotice ale oligodendrogliei si a axonilor. Exista substante blocante a receptorilor alfa-amino-3hidroxi-4metil-4isoxazolepropionic acid, care pot proteja celulele oligodendrogliale si axonii fata de neurotransmitatorii excitatori tip glutamat, dar nu pot bloca raspunsul imun fata de antigenele mielinice. In concluzie, putem spune ca reactiile inflamator-autoimune distrug mielina prin actiunea directa a macrofagelor, dar si prin intermediul autoanticorpilor, citochinelor si complementului.
Dupa cum am mai afirmat, cauza SM este necunoscuta, dar se crede ca implica trei factori: vulnerabilitatea genetica (mostenire de prea multe gene susceptibile si prea putine gene de protectie), vreo forma de expunere la unul sau mai multi agenti patogeni din mediu si dezvoltarea raspunsului imunologic patogen directionat impotriva sistemului nervos central (SNC).
Pana in ultimul deceniu nu a existat tratament pentru SCLEROZA MULTIPLA. Din 1993, cinci terapii de modificare a bolii (DMT) au fost aprobate. Patru sunt imunomodulatoare, iar unul este imunosupresiv. Toate terapiile de modificare produc efecte multiple asupra sistemului imunitar, care ar trebui sa amelioreze SCLEROZA MULTIPLA. In etiopatogenia SM exista cinci caracteristici: 1. natura fundamentala a procesului distinctiv se poate schimba in timp; 2. Implicarea axonului si a neuronilor este o trasatura centrala; 3. Cea mai mare parte a activitatii bolii este inaparenta (subclinica); 4. Exista anomalii microscopice extinse in tesutul creierului cu aparenta normala; 5. SM este eterogena (de natura diferita sau evolutie diferita).
Desi vulnerabilitatea genetica este probabil cea mai importanta pentru dezvoltarea SM, nu este suficienta, in absenta altori factori. Cu un taram recunoscut scazut si zone geografice cu risc ridicat de boala, factorii de mediu joaca in mod clar un rol. Expunerea la virusi si bacterii obisnuiti, destul de timpuriu in viata, intr-un mod care nu este inca inteles pregateste scena pentru SCLEROZA MULTIPLA. Infectiile, probabil actioneaza ca agenti de boala, desi infectia continua activa neurala sau extraneurala nu a fost eliminata ca factor de boala, la anumiti pacienti. Se crede, desi nu s-a dovedit, ca reactivitatea agentului infectios la componenta mielina initiaza SM, la indivizi vulnerabili genetic. Doua tipuri de mecanisme pot fi implicate: unul intermolecular si unul intramolecular, in dezvoltarea atacului imun (VANDERLUGT C. L. si colab., 2002). Acest lucru extinde atacul imun si intensifica si perpetueaza boala autoimuna la un anumit organ. TUOHY V. K. si colab., (1997) numeste aceasta extindere raspandirea epitopa si aceasta are loc la modelele animale de SM, iar datele preliminare indica faptul ca este deasemenea un factor etiopatogenic in SCLEROZA MULTIPLA.
Raspandirea epitopa indreptateste inceperea tratamentului efectiv al SM cat mai devreme posibil (ideal la primul atac), pentru a minimaliza expansiunea si repetarea procesului distructiv. Astfel, terapia incipienta trebuie sa fie cu un spectru larg de imunosuprimare, urmata de terapie de mentinere. Proba ce sustine ca procesul timpuriu al bolii este critic, vine din studiul de istorie naturala al primului atac. La pacientii cu sindrom clinic izolat, numarul si volumul leziunilor creierului in T2 la RMN, sunt cele mai puternice corelari de dizabilitate la varsta de 14 ani, urmate de dezvoltarea leziunii RMN in timpul primilor 5 ani de evolutie a bolii (BREX P. A. si colab., 2002).
Se crede ca SM implica un proces de boala bifazica. Devreme, inflamatia este proeminenta, corespunzand fazei de acutizare si reversie a SCLEROZA MULTIPLA. Mai tarziu exista tranzitie spre o faza neurodegenerativa primara, care coerspunde SM progresiva, cu deficite ireversibile. Desi inflamatia si neurodegenerarea sunt detectate in orice moment, un proces pare a fi dominant. Acest concept este compatibil cu studiile de istorie naturala a SM, deoarece majoritatea pacientilor incep cu boala in faza acuta, dar apoi trec la boala progresiva secundara. Prezenta fazelor SM distincte cere terapia care este potrivita cu natura procesului bolii. In mod clasic, SM e considerata o boala care implica SNC in inflamatie si demielinizare. Este clar din datele directe patologice si cele indirecte neuroimagistice ca ea implica si distrugerea axonilor si neuronilor (CIFELLI A. si colab., 2002; KUHLMAN T. si colab., 2002). Atat densitatea, cat si volumul axonilor se reduc in SM, nu numai in placa, dar si in tesutul SNC cu aparenta normala (TRAPP B. D. si colab., 1998; FERGUSON B. si colab., 1997). Analiza N-acetyl aspartat (NAA), un marcher axon/neuron, masurat de RMN cu spectroscopie, indica faptul ca intreg NAA al creierului este redus, chiar si in SM timpurie (GONEN O. si colab., 2002). Pierderea sau restrangerea axonilor e un contributor major la atrofia creierului si maduvei spinarii. La pacientii cu SM, atrofia SNC este prezenta foarte devreme, chiar la primul atac clinic (CHARD D.T. si colab., 2002; DALTON C. M. si colab., 2002). Importanta distrugerii axonilor in SM nu poate fi exagerata, se crede insa ca este substratul neuroanatomic de dizabilitate permanenta si de progresie a bolii. Distrugerea axonilor reflecta fara indoiala factori multipli. Se disting cel putin cateva din elementele imune si inflamatoare care afecteaza axonii, de cele care distrug mielina. Axonii sunt, ca si canalele ionice si neurofilamentare, tinta unui atac imun direct primar sau secundar. Anticorpii la componentele neurofilamentare, gangliozidele, glucoproteina din mielina oligodendrocitelor au fost legate de SM progresiva (GENAIN CP. si colab., 2002; SILBER E. si colab., 2002; TAYYEBEN SADATIPOUR B. si colab., 1998). Alternativ, distrugerea ar putea reflecta efecte trecatoare secundare de la factorii inflamatorii sau toxici eliberati in mediul intern, prin celule intriseci (microglia, astroglia) sau extrinseci (limfocite, macrofage). Afectarea acuta a axonilor, masurata de expresia protein precursor amyloid se coreleaza cu infiltrarea de celule macrofage si CD8+T. In plus pierderea continua de mielina face rau axonilor (intrerupe transportul axonilor cu stress metabolic crescut asupra neuronilor si axonilor) (HERNDON R. M., 2002). Legatura simbolica intre mielina si axoni inseamna ca insasi demielinizarea este un mecanism de distrugere de axoni. Pierderea de mielina, deasemenea, afecteaza functionarea canalelor de ioni pe suprafata golita a axonilor. Aceste schimburi pot restabili transmiterea nervoasa sau produce distrugerea axonilor (WAXMAN S. G., 2001).
Deoarece insertia de noi canale de sodiu permite transmiterea, aparitia mai rapida si intensificata de astfel de canale este un scop terapeutic dorit. In contrast, canalele de calciu - in mod normal exprimate doar la terminalul axonului presinaptic - poate de asemenea fi inserat in membrana demielinizata. Deoarece inserarea acestor canale duce la distrugerea axonilor (mediati de calpaine) se va dori a se impiedica refacerea lor. Manipularea canalelor ionice poate fi tinta terapeutica viitoare in SCLEROZA MULTIPLA.
Cand mor neuronii, si axonii se pierd. Un studiu recent al unui material de autopsie a creierului a examinat talamusul, o regiune cu materie cenusie bogata in neuroni. Creierul cu SM a prezentat cu 30-35% mai putin neuroni decat tesutul creierului normal (CIFELLI A. si colab., 2002). Masuratorile RMN indica tulburari difuze ale materiei cenusii, chiar la pacientii cu SM timpurie. Semnificatia deplina a implicarii neuronilor in SM este neclara, dar ar putea juca rol in tulburarile cognitive. Faptul ca axonii si neuronii sunt distrusi, cu corelari dovedite de dizabilitate clinica si progresie a bolii SM si posibile corelari cu pierderea cognitiva, indica importanta lor ca tinta terapeutica. Strategiile neuroprotectoare, factorii de crestere neurotrofica, terapiile imunomodulatoare, transtul celulelor si chiar transferul de gene sunt abordari de viitor, pentru a face neuronii si axonii mai putin vulnerabili la leziuni.
Prin definitie, pacientii cu SM recidivanta sunt stabili intre atacurile bolii. Inainte se credea ca SM a intrat in remisie clinica. Azi se stie ca toti pacientii cu SM traiesc boala subclinic continuu, chiar si cand nu au loc schimbari sau inrautatiri in examenul neurologic al pacientului. Studii frecvente de RMN indica faptul ca 80-90% din leziunile creierului sunt asociate cu recidive clinice sau schimbari detectabile la examinare (MILLER D. H. si colab., 1998). La indivizii cu SM netratata, leziunile din creier cresc cu 5-l0% anual (MILLER D. H. si colab., 1998). Atrofierea accelerata a creierului este prezenta chiar timpuriu, la pacientii fara acutizari cu scor KURTZE stabil (KALKERS N. F. si colab., 2002). Este clar, ca criteriile clinice subestimeaza activitatea bolii. Cu exceptia SM benigne, studiile de istorie naturala arata ca toti pacientii netratati dezvolta infirmitate. Mai mult, SM benigna nu a fost niciodata definita si implica probabil cam 5-l0% din pacienti (NOSEWORTHY J. H. si colab., 2000). Intr-un studiu, jumatate din cei diagnosticati cu SM benigna la 10 ani de la debut, aveau infirmitate semnificativa dupa 20 ani de evolutie a bolii (HAWKINS S. A. si colab., 1999).
Observatiile clinice subestimeaza procesul adevarat de distrugere in SCLEROZA MULTIPLA. La fel fac si investigatiile RMN. Tehnicile conventionale RMN detecteaza leziunile hiperdense T2. Tehnicile noi neconventionale pot detecta schimbarile microscopice si fiziologice in tesuturile SNC cu aparenta normala. In creierul pacientilor cu SM, pana la 70% din substanta alba cu aparenta normala poate fi anormala. Urmarirea schimbarilor in tesutul cu aparenta normala, poate fi un marcher important pentru severitatea bolii si raspunsul la terapie.
Intr-o analiza recenta schimbarile NAA anuale in intreg creierul au impartit pacientii recidivanti in trei grupe. Cam 20% din pacienti aveau nivele NAA stabile, 55% prezentau o reducere moderata, 25% prezentau o reducere insemnata in aceste nivele (GONEN O. si colab., 2002). Acestia din urma, cu o distrugere mai mare de axoni au boala mai severa, un prognostic mai rau si vor dezvolta o infirmitate progresiva rapida. Imagistica RMN prin transfer de magnetizatie semnaleaza schimbari in moleculele fluide si fixe in diferite regiuni ale sistemului nervos central. RMN prin transfer de magnetizatie permite masurarea gravitatii leziunilor - cu cat e mai scazut transferul de magnetizatie cu atat e mai mare distrugerea tesutului.
Un concept final al SM este ca boala e eterogena (se exprima diferit). Boala demonstreaza clar variabilitatea clinica, bazata pe subtipuri clinice distincte si pe severitatea bolii. Exista si eterogenitate genetica, bazata pe rasa, pe capacitatea de a produce mielina. Doar 70% din pacienti sufera remielinizarea placilor de SCLEROZA MULTIPLA. Factorii care impiedica remielinizarea la 30% din pacienti nu sunt cunoscuti, desi studiile sugereaza ca prezenta precursorilor oligodendrocitelor, integritatea axonilor, exprimarea moleculelor de pe suprafata axonilor sunt contributorii importanti (CHANG A. si colab., 2002; JOHN G. R. si colab., 2002; WOLSWIJK G. si colab., 2002; KORNEK B. si colab., 2001; LUCCINETTI C.F. si colab., 1999). Studiile lui LUCCINETTI C. F. (2000) sugereaza eterogenitatea imuno-patologica. Rezultatele tuturor studiilor arata patru imunopatologii distincte: tipul 1 cu frecventa de 19% are mecanism mediat macrofagic si in care este prezenta remielinizarea; tipul 2 cu frecventa de 53% mediat de anticorpi si complement cu prezenta remielinizarii; tipul 3 cu frecventa de 26% in care se gaseste moartea apoptotica a oligodendrocitelor, care scad ca numar si in care nu se intalneste remielinizarea si tipul 4 cu frecventa de 2% in care predomina degenerarea oligodendrocitelor in scadere vertiginoasa si in care nu se intalneste remielinizarea. Tipul 3 se exprima clinic cu boala Balo (demielinizare concentrica), iar tipul 4 este corelat cu forma clinica de SM primar progresiva.
Un nou studiu arata ca sync tin - proteina produsa de un virus care a existat in materialul genetic uman pentru mai multi ani - poate fi asociat cu distrugerea mielinei ce se produce in SCLEROZA MULTIPLA. Dr. Joseph Anthony Christopher Power (Universitatea Calgary, Alberta) si colegii au raportat descoperirile, in editia din Octombrie 2004 a Nature Neuroscience. Investigatorii au examinat membrana creierului pentru un numar mic de persoane cu SM si au descoperit mai multa proteina sync tin in membrana celor cu SM, cand au at membrana creierului altor persoane, cu alte boli neurologice sau persoane sanatoase. Deasemenea, ei au mai gasit sync tin in zone unde distrugerea mielinei se petrece activ.
S-a demonstrat ca proteina sync tin induce producerea de oxidanti, molecule, care pot fi toxice pentru unele celule. Cercetatorii au descoperit ca, adaugand proteina sync tin mostrelor de celule producatoare de mielina, duce la moartea acestor celule. In cazul unui cobai, injectarea unui virus producator de sync tin duce la distrugerea mileinei si deteriorarea functiei motorii.
Din cauza cresterii vizibile a oxidantilor, cercetatorii au administrat acid feluric - un antioxidant -, celulelor producatoare de mielina, tratate cu sync tin si cobailor. Acest tratament a redus marcant moartea celulelor producatoare de mielina si a imbunatatit anomaliile motoare ale cobailor tratati cu sync tin.
Intr-un editorial insotitor, Dr. Mark Mattson si Dennis Taub (National Institute on Aging, Baltimore) au descoperit ca, desi acest studiu a gasit o asociere intre sync tin si procesele inflamatorii aparute in SM, nu a stabilit un rol al acestei proteine in procesele bolii. Pentru a stabili o legatura, sunt necesare alte studii, de exemplu, determinarea daca blocarea proteinei sync tin imbunatateste starea cobailor cu SM si celulele imunitare de la oameni reactioneaza la aceasta proteina.
Cu toate acestea, daca rezultatele interesante ale acestui studiu pot fi confirmate si aprofundate, se pot deschide noi cai de investigare in patologia fundamentala a SM si pot sugera piste pentru noi abordari ale tratamentului.
D. Encefalomielita alergica experimentala (EAE) a cainelui, model morfopatologic, clinic si imunochimic pentru SM
Pentru a gasi un model experimental cat mai apropiat de bolile demielinizante umane, cercetatorii au studiat posibilitatea producerii encefalitei alergice experimentale (EAE) pe diferite specii de animale. Astfel EAE a fost produsa la iepuri, cobai, sobolani albi, maimuta, etc, prin administrarea de emulsie de creier, la care s-a adaugat adjuvant FREUD, format din microbacterii omorate si amestecate cu ulei de parafina
MISKOLCZY si colaboratorii (1962) dovedesc actiunea adjuvanta a vaccinului antipertusis, care administrat inaintea amestecului encefalitogen maresc receptivitatea la EAE a animalelor de laborator. Cu aceasta metoda KELEMEN (1968, 1970) reuseste sa provoace la cobai, in mod constant EAE. La iepuri declansarea EAE a fost mai dificila, la fel si la sobolanii albi, iar la soareci boala nu s-a produs. De asemenea si alti cercetatori din intreaga lume reusesc sa produca EAE la diferite animale mici de laborator, dar aici ii amintesc doar pe cei din Romania: BECUS T., 1967, DONA VICTORIA, 1972, DUMA si colaboratorii 1966, FESZT, 1968, G NDISCH, 1966, LAZAR, 1965, MAROS, 1961, PETRESCU, 1967, POPOVICIU, 1967, FLORICA VERDES, 1970, SERBAN, 1970, WAITSUK, 1967, etc.
Majoritatea autorilor considera ca EAE realizata la animalele mici de laborator nu este identica cu encefalomielitele demielinizante umane, neputand fi folosite ca modele experimentale pentru studiul acestora. S-a ivit necesitatea gasirii unui animal de experienta mai apropiat omului, al carui EAE sa poata fi folosit ca model experimental pentru bolile demielinizante umane.
Aproape toate EAE a animalelor mici de laborator sunt din punct de vedere morfologic o encefalita acuta hemoragica. Chiar daca in ultimul timp s-au selectionat genetic o serie de suse de cobai care ar face o EAE foarte asemanatoare SM din punct de vedere morfologic si al evolutiei, noi credem ca din punct de vedere imunologic aceasta nu poate fi ata cu boala la om, intrucat bagajul imunologic al cobaiului si omului se deosebesc destul de mult. Or, tocmai studiile imuno-genetice sunt acelea, care in ultimul timp au revolutionat teoriile privind etiologia si patogenia SCLEROZA MULTIPLA. In cercetari de mai multi ani, MAROS si colaboratorii (1959, 1961, 1964, 1965, 1966, 1968) reusesc sa demonstreze ca EAE a cainelui este cea mai apropiata morfologic encefalomielitelor umane. Cercetari asupra EAE a cainelui s-au facut putine in lume, amintind aici pe SCHERER, (1937), dar acestea n-au sesizat posibilitatea mare de asemanare din punct de vedere imunologic a acesteia cu SM a omului.
Folosind modelul lui MAROS, noi am incercat sa demonstram ca EAE a cainelui este asemanatoare SM, atat din punct de vedere morfoogic, dar mai ales imunologic si clinic. In acest submodul incercam sa demonstram acest lucru:
In cercetarile noastre am folosit 36 de animale impartite in 4 loturi. Lotul M a cuprins 6 animale folosite ca si martori. Lotul A a fost format din 10 animale, pe care dupa producerea EAE s-au studiat modificarile clinice, imunochimice si histomorfologice ale EAE a cainelui. Lotul B a fost format din 10 animale, carora dupa procerea EAE li s-a administrat ACTH, pentru a studia efectul acestui medicament asupra modificarilor clinice, imunochimice si histomorfologice ale EAE a cainelui. Lotul C a fost format din 10 animale, carora dupa producerea EAE li s-a administrat Imuran (azathiopirina), pentru a observa efectul ecestui medicament asupra modificarilor clinice, imunochimice si histomorfologice ale EAE a cainelui.
Pentru producerea EAE la caine am folosit o suspensie cerebrala, preparata dupa reteta profesorului dr. MAROS (1964). Substanta nervoasa bovina, in cantitate de 75 g a fost recoltata de la abator in conditii de sterilitate. Aceasta a fost titrurata in 150 g solutie fenolata 0,25%, suspensiei astfel obtinute adaugandu-i-se 225 g adjuvant Freud. Adjuvantul Freud l-am preparat din 215 ml ulei de parafina, la care am adaugat 4 mg bacili Koch umani omorati prin autoclavare, la fiecare 1 ml de parafina. Astfel, in final, in suspensia noastra, bacilii Koch au fost in concentratie de 2 mg la 1 ml amestec encefalitogen. Suspensiei astfel obtinute i-am adaugat si 10 ml tween. Amestecul encefalitogen se poate pastra bine in sticle de culoare inchisa, la frigider, fara a-si pierde eficacitatea timp de 6 luni. Inainte de folosire, amestecul trebuie agitat si incalzit la temperatura corpilui animalului.
Bazandu-ne pe datele publicate de MAROS (1964,1965), am reusit sa marim eficienta metodei de producere a EAE la caine, administrand doze mici de antigen in regiunea cefei.
Inainte de inceperea administrarii amestecului encefalitogen, de la toate animalele am recoltat sange si LCR pentru determinari imunologice. Tinand cont de cele aratate de MISKOLCZY si colaboratorii (1960,1962), pentru marirea susceptibilitatii alergice a animalelor am administrat intraperitoneal la toate animalele 1 ml de vaccin diftero-tetano-pertusis. La trei zile de la vaccinare, am inceput sa administram cate 1 ml suspensie encefalitogena, injectarea facandu-se la interval de 5 zile in regiunea cefei si a spatelui, intracutan.
Dupa aparitia simptomelor clinice de boala, de la animalele bolnave si chiar de la cele neimbolnavite, am recoltat periodic sange si LCR, pentru investigatii dinamice imunologice.
De asemenea, animalele au fost supravegheate si examinate zilnic pentru descrierea tablourilor clinice a EAE la caine, aparute la animalele noastre.
Animalele din lotul A au fost sacrificate dupa imbolnavire, pentru studiul modificarilor histomorfologice in EAE la caine. Cele din lotul B si C au fost tratate dupa imbolnavire cu ACTH si respectiv cu Imuran, timp de 15 zile. ACTH-ul l-am administrat intramuscular, in doza de 5 u.i. pe Kg.corp/ zi, iar Imuranul l-am administrat per os in doza de 5 mg. pe Kg.corp/zi. La incheierea tratamentului am sacrificat si aceste animale pentru a studia efectul celor doua medicamente asupra leziunilor histomorfologice din EAE la caine. De asemenea am studiat zilnic efectul ACTH-ului si Imuranului asupra simptomelor clinice ale EAE la animalele noastre.
Cainii au fost sacrificati in faza de agravare a bolii, respectiv dupa terminarea tratamentului, la animalele din lotul B si C prin injectarea intracardiaca de 10 ml formalina concentrata, sub narcoza. Dupa sacrificare, am recoltat fragmente de tesut din diferite parti ale SNC, pe care le-am fixat in formalina 10%. Sectiunile la gheata au fost colorate pentru mielina dupa metoda LAZAR (1955, 1960, 1966, 1967). Deasemenea am folosit coloratia cu hematoxilina - eozina, coloratia NISSL si impregnatia argentinica.
1. Tabloul clinic al EAE la caine
Animalele noastre au reactionat foarte variat la amestecul encefalitogen. Astfel, sapte animale au facut boala dupa 12 zile (doua inoculari); patru animale dupa 14 zile (trei inoculari); trei animale dupa 21 de zile (cinci inoculari); noua animale dupa 28-30 de zile (sase inoculari); iar sapte animale nu au prezentat simptome clinice de boala, nici dupa opt saptamani de inoculari.
Cele mai frecvente simptome intalnite au fost: deficitele motorii, constand in tetraplegie, hemiplegie, paraplegie, si diplegie. Au urmat, in ordinea frecventei, tulburarile de coordonare si de echilibru, insotite de tremuraturi cu caracter intentional, toate acestea conturand tabloul clinic al unei ataxii de tip cerebelos. Au urmat apoi: tulburarile de vedere (chiar cecitate), mioclonii localizate sau generalizate, convulsii tonico-clonice, contracturi in extensie a membrelor, uneori hipotonie generalizata si tulburari vegetative, dintre care cel mai des intalnite au fost tulburarile de respiratie si tulburarile sfincteriene.
Animalele care nu au prezentat semne clinice obiective neurologice, aveau totusi unele modificari de comportament ca: irascibilitate, somnolenta, apatie, inapetenta si unele - agresivitate marcata.
Privind intensitatea simptomelor, am avut animale cu forme grave de boala, care au prezentat in general un debut brusc, animale cu forme medii de boala si forme fruste, la care debutul a fost lent insidios. Rezulta ca in experimentul nostru, simptomele cele mai des intalnite au fost: deficitul motor, ataxie, tulburarile de vedere, si cele sfincteriene, simptome care se intalnesc foarte frecvent si in SM (POPOVICIU, 1967, 1969, 1975); CAMPEANU, 1974, SERBAN, 1980). La fel ca si in SM - si in EAE la caine, am observat ca factorii meteorologici au o influenta agravanta. Toate aceste constatari subliniaza marea asemanare clinica dintre EAE la caine si SCLEROZA MULTIPLA.
Pornind de la teoria patogeniei imunoalergice a EAE, am cautat in lucrarea noastra sa studiem efectul unor medicamente antialergice si imunosupresoare asupra simptomelor clinice a EAE la caine, administrand ACTH si Imuran dupa metodologia descrisa mai sus.
In urma administrarii ACTH-ului, simptomele clinice a EAE s-au ameliorat evident, la majoritatea animalelor. De fapt, acest preparat se foloseste cu rezultate bune si in clinica umana, pentru combaterea puseelor acute din SCLEROZA MULTIPLA.
Dupa administrarea Imuranului, la animalele bolnave am constatat o ameliorare rapida a bolii si chiar vindecare clinica, la mai bine de jumatate din animale. Ramane ca si Imuranul sa fie incercat cu mai mult curaj in tratarea SM, respectand un dozaj si o durata de tratament la care sa nu apara unele efecte secundare intalnite la animalele noastre de experienta (accentuarea procesului infectios la nivelul plagilor create la locul inocularii amestecului encefalitogen).
2. Modificarile histomorfologice caracteristice EAE la caine
Studiind preparatele obtinute de la cei 10 caini din lotul A, sacrificati dupa imbolnavire, am observat ca principalele etape de dezvoltare ale modificarilor histo-morfologice in EAE la caine sunt: dilatarea spatiilor perivasculare; aparitia infiltratelor hematogene perivasculare; perivasculita acuta sau subacuta; panvasculita; demielinizari perivasculare strict limitate; contopirea focarelor demielinizate perivasculare; aparitia extravazatelor noi in interiorul sau in jurul focarului demielinizant deja format; edemul perifocal de-a lungul caruia procesul se proa in sens centrifug; aparitia edemului difuz insotit frecvent de extravazare, invazii hematogene, eventuale microhemoragii pe teritoriile interpuse focarelor demielinizante; formarea focarelor de demielinizare difuza. Aceste modificari histo-morfologice au fost gasite la toate nivelele SNC al animalelor bolnave de EAE. Chiar la animalele fara semne clinice evidente de boala, aceste modificari histopatologice s-au intalnit in asa zisele zone mute ale SNC ca lobul frontal si temporal.
Din analiza preparatelor obtinute de la cainii bolnavi de EAE si netratati, rezulta caracterul imunoalergic al procesului de encefalomielita. Focarele infiltrative perivasculare cu tendinta de expansiune centrifuga, caracterul mixt hematomononuclear (foarte variabil), amintesc de leziunile intalnite la om in formele incipiente ale SCLEROZA MULTIPLA.
Mare parte a leziunilor surprinse de noi corespund morfologic unei leucoencefalomielite subacute cu tendinta la cronicizare, fara reactii evidente din partea nevrogliei. De fapt, aceasta este etapa evolutiva in care si la om ne putem astepta la rezultate terapeutice in cazurile de SCLEROZA MULTIPLA. Daca procesul trece in faza de constituire a placilor sclero-nevroglice, evident ca orice tendinta terapeutica este sortita la esec.
De aceea, credem ca tabloul morfologic realizat in experientele de fata poate fi socotit ca un model experimental pentru urmarirea efectelor drogurilor imunosupresive si antialergice, in stadiile incipiente ale SCLEROZA MULTIPLA.
Analiza microscopica a pieselor obtinute de la animalele din lotul tratat cu ACTH si lotul tratat cu Imuran, evidentiaza o actiune antialergica si antiinflamatoare a ambelor preparate.
Astfel, in cazul terapiei cu ACTH, infiltratele celulare perivasculare apar rar, tecile de mielina sunt bine conurate, iar ariile de demielinizare constituie o raritate. La fel si pericarionii au aspect normal, fara nici cele mai discrete alterari; focarele microhemoragice se schiteaza foarte rar.
De asemenea si la animalele tratate cu Imuran lipsesc semnele de perivasculita, alterarea pericarionilor, si a tecilor de mielina. In atie cu lotul tratat cu ACTH, efectele mielino-protectoare ale Imuranului apar mai evidente, deoarece la acest lot nu am observat nici cele mai discrete semne de dezintegrare a mielinei.
Rezulta asadar, ca cele doua substante testate influenteaza sensibil egal (imuranul ceva mai evident), procesul inflamator si demielinizarea perivasculara caracteristica EAE la caine.
3. Modificarile imunochimice in EAE la caine
Baza biologica a cercetarii noastre a constat din cei 30 de caini, impartiti in trei loturi, carora li s-a produs EAE. Martorii au fost aceiasi 30 de caini, carora li s-a prelevat sange si LCR inainte de inocularea amestecului encefalitogen, dupa aparitia semnelor clinice de boala si dupa terminarea tratamentului cu Imuran si ACTH (la cele tratate). Animalelor care nu au facut boala li s-a recoltat sange si LCR periodic, pentru a observa si la ele prezenta sau absenta modificarilor imunologice.
Pentru studiul spectrului proteic al serului si LCR-ului recoltat de la animalele bolnave de EAE netratate si tratate am folosit electroforeza si imunoelectroforeza pe gel de agaroza. Acest studiu l-am facut tot timpul ativ, intre serul si LCR-ul de la animalele sanatoase si cele bolnave, intre serul si lichidul animalelor netratate si cele tratate.
Electroforeza ativa dintre serul animalelor sanatoase si cele bolnave, evidentiaza pe de o parte, cresterea fractiunilor II si IV la animalele bolnave (ceea ce demonstreaza un proces inflamator), si pe de alta parte cresterea fractiunilor VI si VII, ceea ce demonstraza o sinteza de anticorpi).
Pe electroforezele serurilor de la animalele bolnave si tratate cu ACTH si Imuran, am observat o scadere a fractiunilor II, IV, VI, VII (mai evidenta la animalele tratate cu Imuran), ceea ce denota o influienta puternica a acestor medicamente (mai ales a Imuranului) asupra procesului inflamator imunologic.
Imunoelectroforeza serului obtinut de la animalele bolnave, evidentiaza o fractiune prealbuminica neta, neintalnita in serul animalelor sanatoase. Aceasta fractiune prealbuminica este in concentratie mult mai mica, sau a disparut total din serul animalelor bolnave si tratate cu ACTH, respectiv Imuran.
Pentru determinarea imunoglobulinei G din serul si lichidul animalelor am folosit imunodifuziunea radiala simpla (pentru ser) si electroimuno-difuziunea (pentru LCR).
Comparand diametrele individuale ale ringurilor de precipitare de la animalele bolnave de EAE si de la cele sanatoase, se observa ca in serul celor bolnave concentratia imunoglobulinei G este direct proportionala cu gravitatea bolii.
O comportare interesanta am gasit-o la cainii tratati cu Imuran si ACTH, la care nivelul imunoglobulinei G serice scade in unele cazuri sub valori obtinute la cainii sanatosi. Aceasta scadere a imunoglobulinei G la cainii tratati mai ales cu Imuran, atrage atentia asupra pericolului unui tratament prelungit cu acest medicament, fara controlul acestei imunoglobuline.
Cu ajutorul electroimunodifuziunii am constatat in lichidul animalelor sanatoase cantitati foarte mici sau necuantificabile de imunoglobulina G, spre deosebire de animalele bolnave de EAE al caror LCR contine cantitati semnificativ crescute de imunoglobulina G. Aceste cantitati de imunoglobulina G au scazut semnificativ din lichidul animalelor, dupa tratament cu Imuran si ACTH.
Prezenta complexelor imune circulante la animalele bolnave in ser si LCR, reflecta o reactie antigen-anticorp, deci prezenta indirecta a anticorpilor antimielitici in serul si lichidul animalelor bolnave de EAE. Determinarea complexelor imune circulante s-a efectuat dupa metoda descrisa de HASKOVA si colaboratorii, modificata de ABRUDAN si colaboratorii (1980). Valoarea maxima exprimata in unitati, pe care am gasit-o in serul cainilor sanatosi a fost de 50 u.i., cifra considerata ca normala. Valorile peste aceasta cifra le-am considerat patologice. La cainii care au facut EAE, valoarea complexelor imune este net superioara valorilor normale, la animalele cu forme grave de EAE complexele imune atingand cifra de 210 u.i. (MIHANCEA si colaboratorii, 1980).
Complexele imune determinate in serul animalelor tratate cu Imuran si ACTH au avut valori mai mici decat cele obtinute in serul cainilor bolnavi de EAE si netratati, scaderea fiind mai evidenta la lotul tratat cu Imuran.
La putinele cazuri la care am putut doza complexele imune circulante in LCR, am observat ca acestea sunt prezente in cantitati mai mari in lichidul animalelor bolnave, decat la cele sanatoase, precum si scaderea acestor cantitati in lichidul animalelor bolnave, dupa tratamentul cu Imuran si ACTH.
4. Comparatii histopatologice intre EAE a cainelui si SM
a. Studiind caracterul
morfochimic al procesului de demielinizare
din EAE la caine si SM, se constata ca in
ambele afectiuni demielinizarea
comporta in principal modificari fizice si chimice ale
mielinei. Dezintegrarea
fizica, urmata de cea chimica intereseaza toti componentii chimici ai tecii de
mielina. In ambele afectiuni, imaginea histochimica este
dominata de
compusi sudanofili, care dau o reactie pozitiva pentru esterii de colesterol si
care rezulta din descommpunerea lipidelor mielinice. Procesul de
demielinizare din ambele boli este initiat de cresterea activitatii enzimelor
proteolitice si de scaderea proteinei bazice din teaca de mielina.
b. Caracteristic pentru EAE, cat si pentru SM este demielinizarea de
tip primar, in care cel putin in
primele faze, sunt afectate numai tecile de
mielina. Totusi, integritatea prelungirilor cilindraxiale si a celulelor nu
sunt
absolute, in ambele boli.
c. Atat in EAE cat si in SM apar leziuni morfologice si histochimice
ale oligodendrogliei, astrogliei si microgliei, unele diferente privind
intensitatea acestora si perioada cand apar. (MIHANCEA -
1977)
d. Atat in EAE cat si in SM, cea mai caracteristica modificare vasculara
este prezenta infiltratelor perivasculare, cu celule mononucleare de tip limfo-
plasmocitar in spatiile Virchow-Robin si parenchimul
paravascular. In
contextul modificarilor vasculare, atat in EAE cat si in SM s-au pus in
evidenta
tulburari de permeabilitate a barierei hematocerebrale
e. In EAE la caine, leziunile morfopatologice sunt situate mai ales in
substanta alba, mai putin in cea cenusie a SNC, indeosebi in vecinatatea
LCR. In SM, leziunile in aparenta sunt dispuse fara nici o ordine in substanta
alba din intreg SNC, dar mai ales in
apropierea LCR. Nu rareori se constata
leziuni demielinizante si in substanta cenusie
(MlHANCEA - 1977).
f. Privind structura leziunilor demielinizante si stadiul
lor evolutiv in
raport cu durata bolii si momentul observatiei anatomo-patologice, situatia
difera de la EAE la SM in sensul ca:
f1) in EAE leziunile demielinizante se dispun perivenos si sunt in general limitate la teritoriile infiltrate cu celule mononucleare si precipitate plasmatice. In SM focarele de demielinizare sunt izolate sau confluente, dispuse perivenos in forma de placi;
f2) in EAE leziunile demielinizante se gasesc in aceeasi etapa de dezintegrare mielinica, pe cand in SM se remarca stadii evolutive diferite in raport cu momentul observatiei si durata bolii;
f3) leziunile demielinizante din EAE nu prezinta caracterele de progresie periferica intalnite la leziunile din SCLEROZA MULTIPLA.
5. Comparatii imunopatologice intre EAE a cainelui si SM
Intr-un studiu din 1975, MACKAY, CORMEQIE si COATES, stabilesc sase termeni de atie imunologica intre EAE si SM, pe care i-am acceptat si noi, and EAE la caine cu SCLEROZA MULTIPLA.
a. Existenta unui
antigen cunoscut si caracterizat. Se stie ca in EAE
antigenul este cuprins in molecula de proteina bazala a mielinei, cu o
structura cunoscuta. In SM acest antigen inca nu
este cunoscut, existand
mai multe ipoteze cu privire la etiologia bolii (teoria imuno-genetica;
autoimuna).
b. Se pune apoi intrebarea daca exista in aceste boli un raspuns imun
fata de un antigen specific pentru SNC. In EAE, raspunsul este
dirijat
impotriva proteinei bazale a mielinei. In SM,
nu s-au demonstrat inca anticorpi
impotriva unui antigen neuronal sau glial, in
schimb s-au evidentiat
raspunsuri imun-umorale. Indirect prin cresterea titrului de imunoglobuline,
s-a demonstrat existenta anticorpilor umorali antimielinici (POPOVICIU -
1967; BECUS - 1967; SZAB - 1967).
c. Privind existenta
unui raspuns impotriva unui antigen neidentincat,
putem spune ca in EAE nu exista (aici antigenul este cunoscut), pe
cand in
SM exista un asemenea raspuns, manifestandu-se prin nivelul
crescut de
imunoglobulina G in placile scleroase si in
LCR-ul bolnavilor de SCLEROZA MULTIPLA. Acestea
sunt argumente pentru a sustine existenta unui raspuns imun la un antigen
care poate fi extrinsec (viral) sau intrinsec (autoantigen).
d. Existenta unui raspuns imunologic celular impotriva antigenului
specific cerebral apropie mult EAE de SCLEROZA
MULTIPLA. Acest raspuns imun celular a fost
demonstrat atat in EAE cat si in SM cu ajutorul transformarii limfoblastice si
a toxicitatii limfocitelor animalelor bolnave de EAE, respectiv a bolnavilor
de SCLEROZA MULTIPLA.
e. Studiind efectul imunosupresoarelor in EAE si SM, putem afirma
ca EAE este suprimata de aceste medicamente (MlHANCEA si MAROS -
1980). In SM ele suprima puseele acute, boala putand
reveni (POPOVICIU,
1979).
f. EAE poate fi
suprimata prin vaccinari cu doze crescande de proteina
bazica. Si in SM s-au produs diferite vaccinuri (
folosirea copolimerului
1 si proteina bazala orala).
Problema care ramane de rezolvat in continuare, este gasirea antigenului in SM, fata de care apare raspunsul imun umoral si celular. Odata cu gasirea etiologiei SM - si credem ca nu va trece mult pana atunci -medicamentele pentru tratarea acestei boli nu vor intarzia sa apara.
6. Comparatii clinice intre EAE a cainelui si SM
Din punct de vedere clinic, EAE la caine si SM se apropie foarte mult. Atat in EAE, cat si in SM sunt intalnite simptomele: deficite motorii (tetraplegie, paraplegie, hemiplegie, monoplegie, diplegie), ataxia cerebeloasa, tulburari de vedere si tulburari sfincteriene. In ambele afectiuni, factorii meteorologici au o influenta negativa. Evolutia celor doua boli difera una de alta. Daca in EAE simptomele dispar in urma tratamentului, fara sa mai revina (daca nu se administreaza din nou amestec encefalitogen), in SM boala evolueaza in pusee, vindecarea unui puseu neinsemnand vindecare definitiva.
7. Concluzii
Rezultatele obtinute prin cercetarile personale, rezultate care confirma sau infirma unele date din literatura, aduc si date noi in ceea ce priveste modificarile histomorfologice din EAE la caine si posibilitatile de influentare a lor prin unele medicamente. Rezultatele obtinute in cercetarile efectuate pe cel mai la indemana model experimental, mult asemanator SM umane, permit sa tragem urmatoarele concluzii:
a. Modelul nostru experimental nu este costisitor si se afla la indemana tuturor cercetatorilor, care doresc sa studieze problematica bolilor demielinizante.
a. Din cele 30 animale carora li s-a administrat amestec encefalitogen, sapte (25,5%) au facut o forma clinica de boala frusta, la sase animale (20%) s-a manifestat o simptomatologie clinica de forma medie de boala, iar zece animale (55,4%) au prezentat o forma grava de EAE. Sapte animale (25,5%) nu au prezentat simptome clinice obiective de EAE.
b. Perioada care a trecut de la inceperea inocularilor cu amestec
encefalitogen, pana la aparitia primelor simptome clinice de boala,
a variat
de la douasprezece zile (sapte animale adica 25,5%), patrusprezece zile (patru
animale, 15,5%); douazeci si una zile (trei animale, 10%), pana la douazeci
si opt zile (noua animale, 50%).
c. Formele
clinice grave de EAE in general au aparut numai dupa
doua sau trei inoculari, pe cand formele usoare, dupa
cinci sau sase inoculari
de amestec encefalitogen.
d. Cele mai frecvente simptome intalnite la animalele bolnave de EAE
au fost: deficitele motorii (tetraplegie, paraplegie, monoplegie), tulburari de
coordonare si de echilibru, tulburari de vedere, miscari involuntare de tipul
miocloniilor si tremuraturilor, contractura musculara si tulburari vegetative.
Se evidentiaza ca majoritatea acestor simptome se intalnesc si la om, in SCLEROZA
MULTIPLA.
e. Atat
debutul cat si evolutia simptomelor clinice ale EAE la caine au
fost influentate in sens negativ de catre fronturile meteorologice.
Umezeala,
schimbarile bruste de temperatura (mai ales caldura) si infectiile, se stie ca
influenteaza negativ si evolutia SM la om.
f. ACTH-ul, la fel ca in SM,
are influenta favorabila asupra simpto
matologiei clinice si evolutiei EAE la caine, de unde se poate
concluziona
fondul alergic al ambelor boli.
g. Imuranul ofera o buna protectie in EAE
la caine, ameliorand-o,
unerori pana la disparitia simptomelor clinice de boala.
Subliniem ca in
timpul tratamentului cu Imuran am constatat accentuarea procesului infectios
la nivelul plagilor create la locul inocularii amestecului encefalitogen.
Acestea
s-au suprainfectat mai mult ca la animalele netratate cu acest medicament.
Prin urmare, Imuranul trebuie administrat in cure
scurte si combinat cu
medicamente stimulatoare ale hematopoezei. Ramane ca
Imuranul sa fie
incercat mai mult in tratarea SM, respectand un
dozaj si o durata de tratament
la care sa nu apara efectele secundare mentionate mai sus.
h. Alterarile morfologice din SNC al animalelor bolnave, poarta amprenta raspunsului imunoalergic, desfasurandu-se pe un fond vascular compromis, la care se asociaza reactiile secundare de natura inflamatorie si demielinizanta.
i. Substantele cu efect antialergic (ACTH) si imunosupresor (Imuranul), actioneaza spectacular asupra tabloului morfologic al EAE, influentand calitativ si cantitativ substratul lezional al procesului patologic. Efectele se traduc prin suprimarea aproape integrala a reactiilor vasculare si a procesului inflamator asociat, prin protejarea structurilor mielinice si inlaturarea demielinizarii perivasculare, fenomen constant in EAE netratata.
i. Privind modificarile imunochimice intalnite in EAE la caine, putem spune ca separarea electroforetica a proteinelor serice de la cainii bolnavi de EAE ar putea constitui o metoda de monitorizare a diagnosticului si a eficientei tratamentului in timp, desigur daca aceleasi modificari se pot transpune la om.
j. Imunoelectroforeza LCR evidentiaza net existenta arcurilor de precipitare in zonele VI si Vll, care sunt absente la animalele sanatoase sau diminuate (zona VII) la animalele tratate cu Imuran si ACTH.
k. Imunoelectroforeza ativa a serurilor prelevate de la cainii sanatosi, bolnavi tratati si netratati, prezinta doua aspecte care merita retinute: cresterea fractiunilor prealbuminice in cazul animalelor bolnave si netratate, ativ cu aspectul proteic al serului cainilor tratati, la care prealbumina prezinta o concentratie semnificativ mai mica. Concomitent, se observa scaderea albuminei totale la animalele bolnave ativ cu animalele sanatoase.
l. Imunoelectroforeza ativa din LCR si serul aceluiasi animal bolnav de EAE, releva prezenta majoritatii antigenilor serici si in LCR, cu exceptia fractiunii prealbuminice, lucru de altfel demonstrat si prin electroimunodifuziunea practicata cu lichidul concentrat de cinci ori, de la animalele bolnave.
m. Prezenta in LCR a fractiunii prealbuminice, cu diminuarea ei in ser dupa tratamentul cu ACTH, ar putea avea implicatii in aprecierea eficientei acestui tratament. Relevarea acestei observatii ar putea duce la concluzia necesitatii urmaririi acestei fractiuni in lichidul concentrat, pe timpul tratamentului.
n. Electroimunodifuziunea reprezinta un mijloc de investigatie, care alaturi de celelalte investigatii, da posibilitatea monitorizarii eficientei tratamentului in EAE la caine, cu aplicatii in patogenia umana, precum si prognostic asupra evolutiei clinice a bolii.
o. Determinarea imunoglobulinei G in LCR (cu ajutorul electro-imunodifuziunii) si in serul (cu ajutorul imunodifuziunii radiale) animalelor bolnave de EAE, ne-a evidentiat cresterea semnificativa a acestei imunoglobuline, atat in LCR cat si in ser, ceea ce demonstreaza natura imunoalergica a acestei boli.
p. Aceeasi determinare a imunoglobulinei G in LCR si in serul animalelor bolnave de EAE, dupa ce au fost tratate cu ACTH, respectiv cu Imuran, releva ca aceste doua medicamente scad nivelul imunoglobulinei atat in ser cat si in LCR. Scaderi mai semnificative au aparut la lotul tratat cu Imuran.
q. Importanta determinarii imunoglobulinei G in serul cainilor bolnavi de EAE netratati si tratati, rezulta in faptul ca ofera posibilitatea urmaririi tratamentului si intreruperii lui atunci cand concentratia imunoglobulinei G scade sub valori critice.
r. Determinarea complexelor imune aduce un nou mijloc de investigatie, atat in patologia experimentala, cat si in cea clinica umana. Prezenta complexelor imune atesta indirect prezenta anticorpilor antimielinici in serul si lichidul animalelor bolnave de EAE. Semnificatia prezentei anticorpilor antimielinici este foarte bine cunoscuta, dar evidentierea lor in serul si lichidul animalelor bolnave necesita o tehnica foarte laborioasa de imunofluorescenta, care nu este disponibila la nivelul tuturor unitatilor sanitare din tara.
s. La animalele care au facut EAE, valoarea complexelor imune in ser este net superioara valorii normale (50 u.i.), la animalele cu forme grave de EAE valorile complexelor imune atingand cifra de 210 u.
s. La putinele cazuri la care am putut practica determinarea complexelor imune in LCR, in timpul bolii, se demonstreaza prezenta lor in LCR-ul animalelor bolnave de EAE, dar certitudinea o vom putea avea numai repetand investigatiile, dupa unele modificari pe care consideram ca trebuie sa le aducem la recoltarea LCR.
t. Complexele imune determinate in serul si lichidul animalelor tratate cu Imuran si ACTH au avut valori mai mici decat cele obtinute de la cainii bolnavi de EAE si netratati. Desigur, o scadere mai mare a complexelor imune s-ar fi putut obtine daca determinarea lor s-ar fi facut pe un interval de timp mai lung.
t. In patologia umana, determinarea complexelor imune ar putea avea importanta deosebita in urmarirea dinamica a bolnavilor cu SCLEROZA MULTIPLA. Mentinerea unor nivele ridicate in timp, ar putea fi un indiciu de evolutie nefavorabila.
u. Studiind formele clinice de EAE obtinute la animalele noastre si nivelul imunoglobulinei G, precum si al complexelor imune circulante, in ser si LCR, observam ca intre manifestarile clinice si cele imunochimice exista o corelatie directa. Formele grave de EAE au avut semnificativ niveluri ridicate de imunoglobulina G si complexe imune, iar formele mai usoare au prezentat niveluri mai scazute ale imunoglobulinei G si a complexelor imune. Animalele carora li s-a injectat amestec encefalitogen, dar n-au facut boala clinic, nu au avut modificari serice si lichidiene ale imunoglobulinei G si complexelor imune circulante.
v. Corelatia direct proportionala este prezenta si intre modificarile histomorfologice intalnite la animalele cu EAE netratate si nivelurile serice si lichidiene ale imunoglobulinei Q si complexelor imune circulante.
w. Faptul ca imunoglobulina G si complexele imune au scazut dupa tratament cu Imuran, atesta ca acest medicament are o eficienta buna in bolile neurologice alergice experimentale si trebuie incercat cu mai mult curaj si in tratarea afectiunilor neurologice umane cu patogenie autoimuna.
x. Cu ajutorul acestui model experimental am realizat un tablou clinic morfopatologic si imunochimic asemanator fazelor incipiente ale SM la om, remarcand o concordanta aproape perfecta intre cele trei aspecte studiate.
y. Modelul intuit si aplicat de MAROS si colab. (1964, 1965, 1966, 1967, 1968, 1978) este un instrument indispensabil in investigarea experimentala a problematicii largi a encefalomielitei alergice la caine, afectiune cu fond imunoalergic mult asemanatoare clinic, morfologic si imunochimic cu SM la om.
z. Boala experimentala provocata pe aceasta cale la o specie (caine) accesibila si cu parametri biologici apropiati de cei ai omului, prezinta o simptomatologie clinica, morfologica si imunochimica bine conturata, usor interpretabila si masurabila prin mijloacele de care dispunem. Modelul promoveaza dezideratul ca boala sa fie reproductibila in proportia cifrei reprezentative, conditie esentiala a tuturor modelelor experimentale de valoare stiintifica.
 |
 |
 |
 |
La originea bolii figureaza o alimentatie bogata in grasimi de origine animala, un mod de viata stresant (incordare nervoasa, necazuri etc), [...] |
Ce este ateroscleroza si ce consecinte are. Ateroscleroza inseamna infiltrarea peretilor arterelor cu grasimi, dintre care colesterolul este dom [...] |
Aceasta boala este asociata de obicei cu hipertensiunea arteriala. in ateroscleroza, se produce o infiltrare a stratului intim al arterelor, dar si su [...] |
Copyright © 2010 - 2025
: eSanatos.com - Reproducerea, chiar si partiala, a materialelor de pe acest site este interzisa!
Informatiile medicale au scop informativ si educational. Ele nu pot inlocui consultul medicului si nici diagnosticul stabilit in urma investigatiilor si analizelor medicale la un medic specialist.
Termeni si conditii - Confidentialitatea datelor - Contact