1. Elemente de structura a muschiului striat
2. Contractia musculara, etape
3. Manifestarile mecanice ale contractiei musculare
4. Aplicarea principiului I al termodinamicii in cazul contractiei musculare
Musculatura reprezinta un element activ care controleaza deplasarea structurilor osoase si miscarile altor structuri. Miscarile datorate muschilor se bazeaza pe capacitatea fibrelor musculare de a utiliza energia chimica procurata in procesele metabolice, de a se scurta si de a reveni la dimensiunile initiale.
Elemente de
structura a muschiului striat
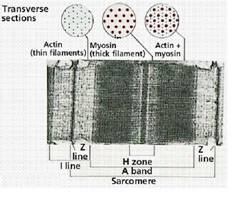
Muschiul striat este alcatuit dintr-un manunchi (sute, chiar mii) de fibre musculare. Fibrele musculare sunt celule polinucleate marginite de o membrana plasmatica - sarcolema, care inveleste fascicule de miofibrile separate intre ele de un sistem de tubuli si cisterne membranoase care apartin reticulului sarcoplasmic. In fibra musculara exista un mare numar de nuclee si organite celulare ca mitocondrii, ribozomi etc. (ura). Unitatea morfofunctionala a miofibrilei este sarcomerul (la homeoterme - lungime 2,2 mm). La microscop sarcomerul apare sub forma unei succesiuni de benzi luminoase si intunecate, care corespund unor filamente longitudinale care se intrepatrund - aceste filamente formeaza aparatul contractil. Sarcomerele sunt separate prin asa-numitele linii Z. Filamentele aparatului contractil sunt de doua feluri: filamente subtiri ( 50) care se insera pe linia Z si filamentele groase ( 100) (ura, ura). Banda luminoasa I contine filamentele subtiri; banda intunecata A contine filamente groase care se intrepatrund cu cele subtiri, iar in centru se afla banda H formata numai din filamente groase. In sectiune transversala se observa urmatoarea imagine: fiecare filament gros este inconjurat de 6 filamente subtiri si fiecare filament subtire de 3 filamente groase.
Filamentele subtiri sunt alcatuite din trei tipuri de proteine: actina globulara (actina G) ( 42.000 D), tropomiozina si troponina. Filamentul propriu-zis este format din doua siruri paralele alipite formate din actina G, ca doua lanturi torsionate (a-helixuri) (150-200 molecule) care dau forma fibrilara (F-actina) (ura).
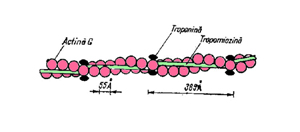
Aceste filamente sunt insotite de cate doua filamente de tropomiozina (70000 D) situate in santurile formate de moleculele de actina. Din loc in loc, la capetele moleculelor de tropomiozina se gasesc moleculele de troponina (50.000 D)(formate din trei proteine). In general, o tropomiozina insoteste 7 molecule de actina.
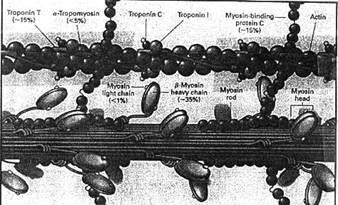
Filamentele groase sunt alcatuite din molecule de miozina (500.000 D) (300-400 molecule) proteina fibrilara, asemanatoare cu o bagheta subtire, terminata prin doua capete globulare (aspect de crosa de hochei) (ura). Partea fibrilara este numita meromiozina usoara (MMU), cea globulara - meromiozina grea(MMG). Aceasta din urma are doua componente: S1 (solubila in apa) si S2 - legatura cu cea usoara; intre ele se poate face indoirea moleculei (ura). Filamentul de miozina este format din portiunea liniara, iar capetele globulare sunt orientate lateral, la exteriorul filamentului (ura), formand asa-numitele punti transversale miozinice (expansiuni laterale). S1 are mare afinitate pentru actina si prezinta, in prezenta Mg++, activitate ATP-azica. La randul ei, actina are o mare afinitate pentru miozina. Prin legarea celor doua (actina si miozina) se formeaza complexul actomiozinic care amplifica activitatea ATP-azica a miozinei de 250 de ori.
Fibra musculara este strabatuta de 2 retele de canalicule: 1) un sistem de canalicule dispuse longitudinal care strabat intreaga sarcoplasma si sunt in contact strans cu miofibrilele: reticulul sarcoplasmic. Acesta se dilata la nivelul liniei Z formand cisternele terminale (ura); 2) canalicule transversale (tubulii T) de la nivelul miofibrilelor pana la suprafata celulei, unde se deschid printr-un por. Acestia se afla in vecinatatea liniei Z. Cisternele reticulului sarcoplasmic si cu tubulii alcatuiesc o triada. Cisternele reprezinta rezervoare de ioni de Ca++ (concentratia de 1000 ori mai mare decat in exteriorul lor )(contin cule in care concentratia ionilor de calciu este de cateva ori mai mare decat in structurile vecine) si acestia se elibereaza in cursul contractiei musculare.
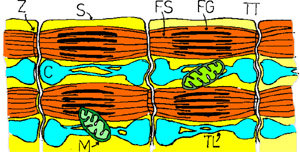
Muschiul relaxat
In repaus, concentratia sarcoplasmica a ionilor de Ca++ este foarte mica ( 10-7 M). In aceasta situatie locurile de legare ale actinei cu miozina sunt mascate prin interpunerea moleculelor de tropomiozina. Puntile transversale MMG nu sunt legate de actina. Deoarece miozina are activitate ATP-azica, poate sa scindeze ATP in ADP si Pi (ura), cu un turnover foarte scazut ( 25 s sau 2-3/min.) si produsii de scindare sunt eliberati in sarcoplasma.
Contractia musculara
Contractia fibrei musculare implica trei etape succesive:
1. excitatia fibrei;
2. cuplajul excitatie contractie;
3. contractia propriu-zisa a fibrei.
1. Excitatia. Nervul motor mielinizat are ramuri terminale nemielinizate. Acestea parcurg o formatiune specializata a sarcolemei formand sinapse neuro-musculare (placa motorie)(ura). La nivelul acestor sinapse incepe excitatia fibrei musculare prin eliberarea moleculelor de mediator chimic (acetilcolina) din terminatia nervoasa in urma unui impuls nervos. Moleculele de acetilcolina se fixeaza pe moleculele receptoare ale membranei postsinaptice, determinand deschiderea unor canale cationice ale acesteia. Cationii intra in fibra, interiorul acesteia devine local pozitiv si in felul acesta se produce potentialul de actiune. Acesta se deplaseaza prin sarcolema in lungul fibrei, iar prin membrana tubilor transversali in profunzime.
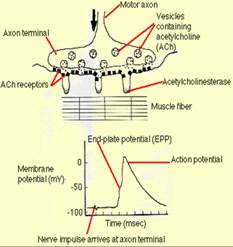
2. Cuplajul excitatie- contractie. Tubulii transversali sunt in legatura cu cisternele reticolului sarcoplasmic. Depolarizarea tubulilor duce, in momentul in care potentialul de actiune ajunge in dreptul cisternelor, la deschiderea canalelor de Ca. Ionii de Ca++ sunt eliberati din cisterne, iar concentratia lor in sarcoplasma creste de la cca 0,1 M la 10 M (100 ori). Troponina fixeaza ionii de Ca si in urma unei modificari conformationale deplaseaza moleculele de tropomiozina din santurile filamentului subtire astfel incat locurile de legare ale actinei cu miozina nu mai sunt mascate (ura).
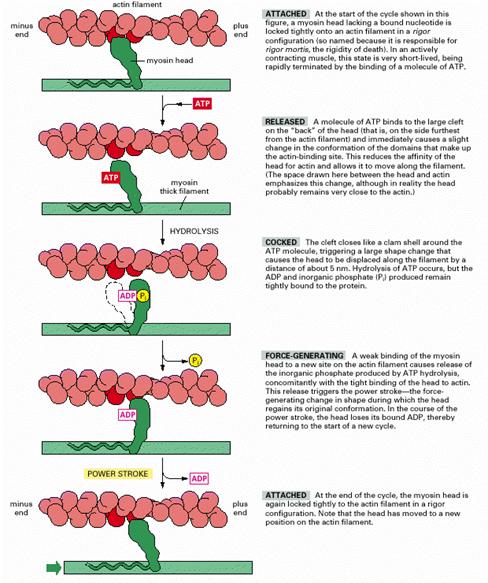
3. Contractia. Din acest moment se poate forma complexul actomiozinic si contractia incepe. Contractia implica eliberarea energiei chimice necesare si fenomenele mecanice care stau la baza producerii fortei, respectiv scurtarii fibrei. Etapele ei sunt:
- In urma rearanjarii troponinei si tropomiozinei extremitatea globulara S1 a miozinei care dispune de 2 locuri de legare, unul pentru actina, altul pentru ATP, se fixeaza pe locul de legare actinic. Dupa cum am spus mai inainte, miozina scindeaza ATP in ADP si Pi si in repaus, dar eliberarea lor este foarte lenta. Complexul ATP- miozina are mare afinitate pentru G-actina.
- Prin formarea complexului actomiozinic activitatea ATP-azica creste de 250 ori, produsii de scindare sunt eliberati rapid, se elibereaza energie, eliberarea Pi induce o modificare conformationala a S2, iar puntea (in pozitie perpendiculara atunci cand produsii de scindare a ATP nu au fost inca eliberati) se inclina cu 450 fata de filament (ura, Alberts et al., 2002). Prin aceasta inclinare ea antreneaza filamentul subtire, deplasandu-l axial cu 7,5 nm in directia zonei mediane si sarcomerul este scurtat. Se dezvolta forta.
- Locul de legare ramanand liber, o noua molecula de ATP este atasata (dupa 10-3 s) si aceasta comanda desprinderea puntii de filamentul subtire (ura). ATP este din nou scindat, puntea se leaga din nou de filament ntr-o pozitie apropiata de linia Z si procesul se repeta.
In felul acesta, puntile transversale se comporta ca vaslele unei barci si sarcomerul se scurteaza (ura, ura). Hidroliza ATP este mai rapida ca in repaus. Turnoverul creste la cca. 10/s. In felul acesta ciclul biochimic al miozinei este continuat, cand incepe contractia, de cel actomiozinic (ura). Observatie: In timpul scurtarii sarcomerelor nu se scurteaza filamentele - ele se intrepatrund - gliseaza. Teoria glisarii a fost elaborata in mod independent de catre A. F. Huxley si H.E. Huxley in 1954.
In lipsa ATP, cum se intampla dupa moarte, ciclul se intrerupe in faza in care puntea este legata de filamentul subtire, fiind inclinata la 450, legatura actomiozinica devenind permanenta. Apare rigor mortis (rigiditatea cadaverica).
Relaxarea
Dupa incetarea excitatiei se produce relaxarea in urmatoarele etape: - se inchid canalele de Ca++ din cisterne, se reface concentratia de 103 ori mai mare in cisterne decat in sarcoplasma, datorita pompei de Ca. Prin scaderea concentratiei calciului, proteinele filamentelor subtiri (troponina) isi reiau conformatia, tropomiozina mascheaza locurile de legare. Atat contractia cat si mentinerea starii de repaus se fac cu consum de energie. Exista muschi ale caror sisteme contractile sunt reglate de ionii de Ca++ prin actiunea acestora asupra miozinei si nu prin intermediul sistemului troponina- tropomiozina, asa cum exista si muschi care pot apela la ambele sisteme de reglare a functiei contractile.
Mecanismele de refacere a rezervei de ATP
Rezervele de ATP din muschi se pot reface pe cateva cai:
Glicoliza anaeroba
Fosforilare oxidativa
Reactia creatin-chinazei (reactia Lohmann):
ADP + CP ATP + C
Reactia miochinazei:
ADP + ADP ATP + AMP
Muschiul glicerinat
Fibra musculara tratata cu solutie de glicerina - devine partial (sau chiar total) lipsita de sarcolema, tubi transversali si reticul sarcoplasmic (o asemenea stare poate fi creata si prin tratare cu detergenti sau prin microdisectie). Se constata ca si in asemenea fibre se pot desfasura cele 2 cicluri daca se modifica concentratia de ioni de Ca++ si ATP in solutia in care se afla fibra. In solutie lipsita de ATP -rigor mortis. La adaugare de ATP - relaxare. La adaugarea ionilor de Ca++ se produce contractia. La indepartarea ionilor de Ca++ (cu EDTA, EGTA -chelatori) se produce ori relaxarea, ori rigor (depinzand de prezenta ATP).
Manifestarile mecanice ale contractiei musculare
Tipuri de contractie
Muschiul dezvolta o forta de contractie egala si de sens contrar fortei careia i se opune. In functie de marimea acestei forte muschiul se poate scurta, alungi sau poate pastra aceeasi lungime.
- Contractie izotonica - muschiul se contracta contra unei forte exterioare constante (ridicarea unei greutati). - Contractie neizotonica - forta variaza ca marime - intinderea unui resort.
- Contractie izometrica - contractie in care lungimea muschiului nu se modifica, dar tensiunea in el creste. Forta dezvoltata este egala cu cea care trebuie invinsa (contractia posturala sau pentru sustinerea unui obiect). Muschiul nu efectueaza lucru mecanic.
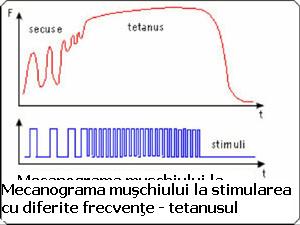
- Contractia tetanica - Prin stimulare cu un impuls unic muschiul se contracta sub forma unei secuse unice (intervalul intre stimuli trebuie sa fie mai lung decat timpul necesar contractiei si relaxarii); la stimulare repetitiva cu o anumita frecventa, peste o limita data, contractiile individuale fuzioneaza intr-o contractie unica - contractie tetanica (ura). Frecventa depinde de tipul de muschi (mai mare la muschii rapizi)(musculatura oculara 350 stimuli/secunda, muschi solear 30).
- Alungirea muschiului - daca forta exterioara este mai mare decat valoarea maxima a fortei pe care o poate dezvolta muschiul, acesta se alungeste cu toate ca se contracta activ.
Relatia forta-lungime
Forta generata intr-o contractie musculara depinde de distanta dintre extremitatile sale - deci de lungimea sarcomerelor sale. Ea are valoarea maxima pentru o lungime initiala a fibrei (in repaus). Daca este intinsa peste aceasta lungime si determinata sa se contracte, sau contractia porneste de la o lungime mai mica, forta de contractie descreste (ura, ura).

Pentru a explica aceasta dependenta trebuie sa se tina cont de numarul de punti transversale implicate in contractie (ura). Daca un muschi este intins peste o lungime a sarcomerelor de 3,6 mm, nu mai are loc intrepatrunderea filamentelor groase si subtiri, deci nu se poate forma complexul actomiozinic si forta va fi nula. Forta maxima atunci cand toate puntile sunt implicate in interactiunea actomiozinica. La muschiul scurtat apare o interpatrundere suplimentara a filamentelor subtiri apartinand aceluiasi sarcomer repezentand un obstacol in interactiunea actomiozinica. F =0 cand filamentul subtire atinge linia Z din partea opusa.
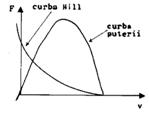
Relatia forta-viteza de scurtare (A.V.Hill)
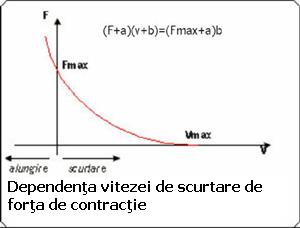
Viteza de scurtare a unui muschi depinde de forta exterioara ce trebuie invinsa (deci de forta de contractie la un moment dat). Astfel, un obiect usor este ridicat mai rapid, unul mai greu, mai lent. Ecuatia care reprezinta acest proces, ecuatia Hill:
(F+a)(v+b) = (Fmax +a)b
Aceasta este ecuatia unei hiperbole (ura). F- forta de contractie, v-viteza de contractie, a,b, -constante (a-dimensiune de forta, b de viteza). Fmax - forta maxima dezvoltata de muschi pentru o anumita lungime initiala la care se declanseaza contractia (izometrica).
De aici, viteza de contractie va fi:
v = (Fmax -F)b/(F+a)
Se obtine viteza maxima, vmax, pentru F=0.
La forte mari muschiul se alungeste si nu mai respecta relatia Hill.
Puterea dezvoltata de muschi
P = Fv = F(Fmax -F)b/(F+a)
Puterea este nula pentru F = 0 si F = Fmax (izometrica) (ura). Ecuatia Hill corespunde unei curgeri vascoase (frecarea la nivelul puntilor in glisare).
Aplicarea principiului I in cazul contractiei musculare
La scurtarea unui muschi cu Dx, impotriva unei forte F, se efectueaza un lucru mecanic L = FDx.
Atat in repaus, cat si in contractie, muschiul degaja caldura. Caldura disipata (ura) de muschi este :
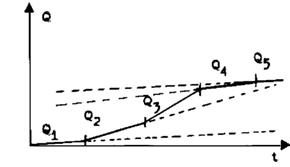
- Q1 - caldura de repaus;
- caldura de contractie, care la randul ei este caldura de mentinere a fortei de contractie Q2 si caldura de scurtare Q3 (proportionala cu gradul scurtarii);
- caldura de relaxare Q4, care se produce imediat dupa incetarea stimularii;
- caldura de restabilire Q5, care se produce in urma reactiilor chimice de regenerare a ATP. Intrucat pentru acest din urma proces este necesar un timp mult mai lung, el nu apare reprezentat in grafic.
Caldura degajata de muschi este rezultatul reactiilor chimice si proceselor mecanice (intinderea unor structuri elastice din muschi, frecari interne). Energia interna a muschiului este rezultatul unor reactii chimice (hidroliza ATP, reactia creatinchinazei, a miochinazei etc.) Variatia energiei chimice se calculeaza pentru fiecare reactie :
DU = xiDUmol i = Q - FDx
unde: xi - numarul de moli de reactanti consumati (gradul de avansare a reactiei)
DUmol i - energia molara a reactiei respective, luata din tabele.
Aspirina are un efect bun; se pot lua 2 tablete pe zi, dupa mese, daca greutatea este sub 75 kg, sau 3 tablete, daca greutatea este mai mare, timp [...] |
BIOFIZICA CONTRACTIEI MUSCULARE 1. Elemente de structura a muschiului striat 2. Contractia musculara, etape 3. Manifesta [...] |
MUSCHII REGIUNII MEDIANE A GATULUI .37 MUSCHII REGIUNII MEDIANE A GATULUI - vedere po [...] |
Copyright © 2010 - 2025
: eSanatos.com - Reproducerea, chiar si partiala, a materialelor de pe acest site este interzisa!
Informatiile medicale au scop informativ si educational. Ele nu pot inlocui consultul medicului si nici diagnosticul stabilit in urma investigatiilor si analizelor medicale la un medic specialist.
Termeni si conditii - Confidentialitatea datelor - Contact
Despre sistemul muscular |
| Alte sectiuni |
|
Ai o problema medicala? |